Spéciation
La spéciation concerne la façon dont les espèces se forment. C'est une partie importante de la biologie de l'évolution. Darwin pensait que la plupart des espèces étaient directement issues d'espèces préexistantes. C'est ce qu'on appelle l'anagén…
La spéciation concerne la façon dont les espèces se forment. C'est une partie importante de la biologie de l'évolution.
Darwin pensait que la plupart des espèces étaient directement issues d'espèces préexistantes. C'est ce qu'on appelle l'anagénèse : les espèces changent, ou "évolution phylétique". Pendant une grande partie du XXe siècle, nous pensions que la plupart des espèces étaient issues d'une division précédente des espèces : la cladogenèse. L'opinion générale était que la plupart des scissions d'espèces sont causées ou favorisées par des mécanismes d'isolement.
La séparation physique des espèces qui ont vécu ensemble est sans aucun doute un facteur essentiel. Elle est illustrée par de nombreux exemples, dont certains sont examinés ci-dessous.
Cependant, les travaux des 20 dernières années ont mis en évidence d'autres causes. L'analyse de la séquence d'ADN des êtres vivants a montré qu'il y a souvent une certaine hybridation entre des espèces apparentées. Cela signifie que des gènes ont été transférés par ces croisements. Cela signifie que l'isolement reproductif n'est pas la seule définition d'une espèce, et que la spéciation ne nécessite pas toujours l'allopatrie (espèces à séparer sur le plan reproductif). Les sections ci-dessous illustrent l'idée que la séparation physique était de première importance dans la formation de nouvelles espèces.
Galerie d’images
10 Images









Mécanismes d'isolement
Les mécanismes d'isolement sont des choses qui empêchent la reproduction réussie entre les groupes d'une espèce. L'isolement reproductif des populations est établi. Ceci est particulièrement important pour le concept d'espèce biologique, car les espèces sont définies par l'isolement reproductif.
Les mécanismes d'isolement peuvent être divisés en deux groupes, avant et après la fertilisation.
Avant la fertilisation
Les facteurs qui empêchent les individus de s'accoupler.
- L'isolement géographique : Les espèces sont présentes dans différentes régions et sont souvent séparées par des barrières.
- Isolement temporel : Les individus ne s'accouplent pas parce qu'ils sont actifs à des moments différents. Il peut s'agir de différents moments de la journée ou de différentes saisons. Les périodes d'accouplement des espèces peuvent ne pas correspondre. Les individus ne se rencontrent pas pendant leurs périodes d'accouplement, ou pas du tout.
- Isolement écologique : Les individus ne s'accouplent que dans leur habitat préféré. Ils ne rencontrent pas d'individus d'autres espèces ayant des préférences écologiques différentes.
- Isolement comportemental : Des individus d'espèces différentes peuvent se rencontrer, mais on ne reconnaît pas les indices sexuels qui peuvent être donnés. Dans la plupart des cas, un individu choisit un membre de sa propre espèce.
- Isolation mécanique : La copulation peut être tentée, mais le transfert de sperme n'a pas lieu. Les individus peuvent être incompatibles en raison de leur taille ou de leur morphologie.
- L'incompatibilité de jeu : Le transfert de sperme a lieu, mais l'ovule n'est pas fécondé.
Après la fertilisation
Les facteurs qui empêchent la réussite de l'accouplement, tels que l'incompatibilité génétique, l'inviabilité des hybrides ou la stérilité.
- Mortalité zygotique : L'ovule est fécondé, mais le zygote ne se développe pas.
- L'inviabilité des hybrides : l'embryon hybride se forme, mais n'est pas viable.
- Stérilité hybride : l'hybride est viable, mais l'adulte qui en résulte est stérile.
- La panne hybride : Les hybrides de première génération (F1) sont viables et fertiles, mais les générations hybrides suivantes (F2 et rétrocroisements) sont inviables ou stériles.

L'isolement géographique
On pense que c'est la cause la plus fréquente de spéciation. La première personne à y penser a été Moritz Wagner, un explorateur allemand et historien de la nature.
Au début de sa carrière, Wagner était géographe et il a publié un certain nombre de livres de géographie sur l'Afrique du Nord, le Moyen-Orient et l'Amérique tropicale. Il était également un naturaliste et un collectionneur passionné, et c'est pour cet ouvrage qu'il est le plus connu des biologistes. Ernst Mayr, évolutionniste et historien de la biologie, a rendu compte de l'importance de Wagner. p562-565
Pendant ses trois années en Algérie, Wagner a étudié (entre autres activités) les coléoptères incapables de voler Pimelia et Melasoma. Chaque genre est divisé en plusieurs espèces, chacune étant confinée à une partie de la côte nord entre les rivières qui descendent des montagnes de l'Atlas vers la Méditerranée. Dès que l'on traverse une rivière, une espèce différente mais étroitement apparentée apparaît.
"... une espèce naissante ne [surgit] que lorsque quelques individus franchissent les limites de leur aire de répartition... la formation d'une nouvelle race ne réussira jamais... sans une longue séparation continue des colons des autres membres de leur espèce".
Il s'agissait d'une description précoce d'un processus d'un type de spéciation géographique. En 1942, elle a été réintroduite par Mayr, et l'importance de la spéciation géographique est devenue l'une des idées centrales de la synthèse évolutionnaire.
Un autre terme de spéciation géographique est la spéciation allopatrique. Allopatrie signifie "terre différente".
Les îles vacantes
Les îles volcaniques sont formées sans vie, et toute vie doit arriver portée par le vent ou l'eau. Nous savons, grâce aux îles hawaïennes et aux îles Galapagos, que toutes les formes de vie changent lorsqu'elles atteignent les îles depuis le continent.
Sur environ 17 000 km2, les îles hawaïennes possèdent la collection de drosophiles la plus diversifiée au monde, vivant des forêts tropicales aux prairies de montagne. Environ 800 espèces de drosophiles hawaïennes sont connues.
Des études montrent un "flux" clair d'espèces des îles les plus anciennes vers les plus récentes. Il y a également des cas de colonisation vers des îles plus anciennes et de disparition d'îles, mais ces cas sont beaucoup moins fréquents.
D'après la datation radioactive au potassium/argon, les îles actuelles datent de 0,4 million d'années (mya) (Mauna Kea) à 10mya (Necker). Le plus ancien membre de l'archipel hawaïen encore au-dessus de la mer est l'atoll de Kure, qui peut être daté de 30 mya.
L'archipel lui-même, produit par le déplacement de la plaque du Pacifique sur un point chaud, existe depuis bien plus longtemps, du moins jusqu'au Crétacé. Les îles hawaïennes et les anciennes îles qui sont maintenant sous la mer forment la chaîne de montagnes sous-marines Hawaï-Emperor ; et beaucoup de montagnes sous-marines sont des guyots.
Toutes les espèces de drosophiles indigènes que l'on trouve sur Hawaiʻi semblent descendre d'une seule espèce ancestrale qui a colonisé les îles, il y a environ 20 millions d'années. La radiation adaptative qui a suivi a été stimulée par un manque de concurrence et une grande variété de niches vacantes. Bien qu'il soit possible pour une seule femelle enceinte de coloniser une île, il est plus probable qu'il s'agisse d'un groupe de la même espèce.
Il y a d'autres animaux et plantes sur l'archipel hawaïen qui ont subi des radiations adaptatives similaires, bien que moins spectaculaires.
Espèces d'anneaux
En biologie, une espèce annulaire est une série de populations voisines reliées entre elles, chacune d'entre elles pouvant se croiser avec les populations voisines. Les deux extrémités de la chaîne se chevauchent.
Les deux populations finales de la série sont trop éloignées l'une de l'autre pour être liées à des croisements. De telles populations "finales" non liées à la race mais génétiquement reliées peuvent coexister dans la même région, fermant ainsi un "cercle".
Les espèces annulaires fournissent des preuves importantes de l'évolution : elles illustrent ce qui se passe au fil du temps lorsque les populations divergent génétiquement. Richard Dawkins a observé que les espèces à anneaux "nous montrent seulement dans la dimension spatiale quelque chose qui doit toujours se produire dans la dimension temporelle".
Il est cependant difficile de trouver un exemple simple et direct.
Mouettes de Larus
Un exemple classique d'espèce de bague est la bague des espèces circumpolaires des mouettes de Larus. L'aire de répartition de ces goélands forme un anneau autour du pôle Nord, qui n'est normalement pas survolé par les goélands individuellement.
En gros, les mouettes peuvent se croiser dans une certaine mesure avec leurs voisines, sauf aux deux extrémités de l'anneau.
Les goélands marins et les goélands argentés sont suffisamment différents pour ne pas s'hybrider normalement ; le groupe de goélands forme donc un continuum, sauf là où les deux lignées se rencontrent en Europe.
Bien que la réalité soit beaucoup plus compliquée, elle montre bien l'idée générale.
Salamandres de l'Ensatina
La salamandre de l'Ensatina est une espèce en anneau dans les montagnes autour de la vallée centrale californienne. Le complexe forme une forme de fer à cheval autour des montagnes. Bien que des croisements puissent avoir lieu entre chacune des 19 populations autour du fer à cheval, la sous-espèce Ensatina eschscholtzii à l'extrémité ouest du fer à cheval ne peut pas se croiser avec l'Ensatina klauberi à l'extrémité est. Il s'agit d'une illustration de "presque toutes les étapes d'un processus de spéciation" (Dobzhansky). Richard Highton a fait valoir que l'Ensatina est un cas d'espèces multiples et non un continuum d'une seule espèce.
La fauvette verdâtre
La fauvette verte (Phylloscopus trochiloides) compte un certain nombre de sous-espèces, dont P. t . viridianus est la plus connue en Europe. Il s'agit d'une espèce en anneau dont les populations divergent à l'est et à l'ouest du plateau tibétain, pour se rencontrer plus tard sur le côté nord. Leurs relations sont assez confuses.




La spéciation sympatrique
La spéciation sympatrique désigne la formation de deux ou plusieurs espèces descendantes d'une seule espèce ancestrale, toutes occupant le même emplacement géographique. On pense aujourd'hui que cette pratique est très courante.
Dans la spéciation sympatrique, les espèces divergent tout en habitant le même endroit. Des exemples souvent cités de spéciation sympatrique se retrouvent chez les insectes qui deviennent dépendants de différentes plantes hôtes dans la même région.
L'existence de la spéciation sympatrique en tant que mécanisme de spéciation a été vivement contestée. On a fait valoir que les preuves de la spéciation sympatrique sont en fait des exemples de spéciation micro-géographique. En général, ce n'est pas l'explication préférée aujourd'hui. Elle est plutôt le résultat d'une hybridation entre des espèces étroitement liées, suivie d'une sélection naturelle travaillant sur la progéniture de ces croisements. Un exemple largement accepté de spéciation sympatrique est celui des cichlidés du lac Nabugabo en Afrique de l'Est, dont on pense qu'il est dû à la sélection sexuelle.
La spéciation par polyploïdisation
La polyploïdie a provoqué de nombreux événements de spéciation rapide parce que les descendants, par exemple, d'accouplements tétraploïdes x diploïdes donnent souvent lieu à une progéniture stérile triploïde.
Cependant, tous les polyploïdes ne sont pas isolés de leurs plantes parentales sur le plan reproductif, et le flux de gènes peut encore se produire, par exemple par des accouplements triploïdes hybrides x diploïdes qui produisent des tétraploïdes.
De nombreuses espèces végétales et la plupart des espèces animales existantes ont apparemment subi une polyploïdisation au cours de leur histoire évolutive. La reproduction des espèces polyploïdes réussies est parfois asexuée, par parthénogenèse. Pour des raisons inconnues, de nombreux organismes asexués sont polyploïdes.
Mouche de l'aubépine
Un exemple d'évolution à l'œuvre est le cas de la mouche de l'aubépine, Rhagoletis pomonella, qui semble subir une spéciation sympatrique.
Les différentes populations d'aubépine se nourrissent de différents fruits. Une population distincte est apparue en Amérique du Nord au XIXe siècle, quelque temps après l'introduction des pommes, une espèce non indigène. Cette population qui se nourrit de pommes se nourrit normalement uniquement de pommes et non du fruit historiquement préféré des aubépines. La population actuelle qui se nourrit d'aubépines ne se nourrit normalement pas de pommes.
Certains indices suggèrent qu'une spéciation sympathique est en train de se produire. Les mouches de l'aubépine arrivent à maturité plus tard dans la saison et prennent plus de temps que les mouches de la pomme ; et il y a peu de preuves de croisement (les chercheurs ont documenté un taux d'hybridation de 4 à 6 %).
L'émergence de la nouvelle mouche de l'aubépine est un exemple d'évolution en cours.
Hybridisation
Rarement, une nouvelle espèce se forme lorsque des membres individuels de différentes espèces s'accouplent. Généralement, les produits de ces accouplements hybrides sont stériles (non fertiles), ou relativement stériles, et sont donc éliminés par la sélection naturelle.
Un exemple d'une nouvelle espèce hybride réussie a été trouvé. La nouvelle espèce est un hybride des moineaux italiens et espagnols, et les faits de base ont été vérifiés par l'analyse de la séquence de l'ADN dans leur sang. L'oiseau vit en Italie dans une région où vivent les deux espèces parentes. Il ne se reproduit pas avec le moineau espagnol, même s'il vit avec eux côte à côte.
La spéciation artificielle
De nouvelles espèces ont été créées par l'élevage d'animaux domestiqués, mais les dates et les méthodes initiales ne sont pas claires. Par exemple, les moutons domestiques ont été créés par hybridation, et ne produisent plus de descendance viable avec l'Ovis orientalis, une espèce dont ils sont issus.
Le bétail domestique, en revanche, peut être considéré comme la même espèce que plusieurs variétés de bœufs sauvages, de gaurs, de yaks, etc., car il produit facilement une progéniture fertile avec eux.
Espèces de laboratoire
Les créations les mieux documentées de nouvelles espèces en laboratoire ont été réalisées à la fin des années 1980. William Rice et G.W. Salt ont élevé des mouches à fruits, Drosophila melanogaster, en utilisant un labyrinthe avec trois choix différents d'habitat tels que clair/foncé et humide/sec.
Chaque génération a été placée dans le labyrinthe et les groupes de mouches qui sortaient de deux des huit sorties ont été mis à part pour se reproduire les uns avec les autres dans leurs groupes respectifs. Après trente-cinq générations, les deux groupes et leur progéniture ont été isolés sur le plan reproductif en raison de leurs fortes préférences en matière d'habitat : ils ne se sont accouplés que dans les zones qu'ils préféraient, et n'ont donc pas pu s'accoupler avec les mouches qui préféraient les autres zones. L'histoire de ces tentatives est décrite dans Rice et Hostert (1993).
Diane Dodd a également pu montrer comment l'isolement reproductif peut se développer à partir des préférences d'accouplement chez Drosophila pseudoobscura après seulement huit générations en utilisant différents types d'aliments, l'amidon et le maltose.
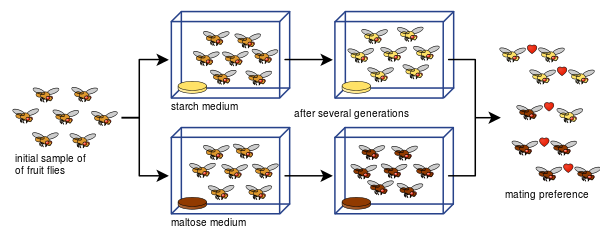
L'expérience de Dodd a été facile à reproduire pour beaucoup d'autres, y compris avec d'autres types de mouches à fruits et d'aliments.
Renforcement (effet Wallace)
Le renforcement est le processus par lequel la sélection naturelle augmente l'isolement reproductif.
Elle peut se produire après que deux populations de la même espèce se soient séparées puis soient revenues en contact. Si leur isolement reproductif était complet, alors elles se seraient déjà développées en deux espèces distinctes et incompatibles.
Si leur isolement reproductif est incomplet, la poursuite de l'accouplement entre les populations produira des hybrides, qui pourront ou non être fertiles. Si les hybrides sont stériles, ou fertiles mais moins en forme que leurs ancêtres, il n'y aura plus d'isolement reproductif et la spéciation aura essentiellement eu lieu (comme chez les chevaux et les ânes).
Le raisonnement est que si les parents de la progéniture hybride ont chacun naturellement sélectionné des traits pour leurs propres environnements différents, la progéniture hybride portera des traits des deux, et ne s'adaptera pas aussi bien à l'une ou l'autre niche qu'à l'autre parent. La faible adéquation des hybrides ferait que la sélection favoriserait les accouplements assortis, ce qui réduirait l'hybridation.
On l'appelle parfois l'effet Wallace, d'après le biologiste évolutionniste Alfred Russel Wallace qui a suggéré à la fin du XIXe siècle qu'il pourrait être un facteur important de spéciation.
Si la progéniture hybride est plus en forme que ses ancêtres, les populations vont alors se regrouper au sein de la même espèce dans la zone où elles sont en contact.
Un renforcement est nécessaire pour la séparation lorsqu'il existe une "zone hybride" entre deux formes d'une espèce. Les zones hybrides sont des régions où des populations divergentes se rencontrent et se croisent. La progéniture hybride est très courante dans ces régions, qui sont généralement créées par des espèces divergentes entrant en contact secondaire.
Sans renforcement, les deux populations ou espèces auraient des croisements incontrôlables. Le renforcement peut être induit dans le cadre d'expériences de sélection artificielle comme décrit ci-dessus.
Questions et réponses
Q : Qu'est-ce que la spéciation ?
R : La spéciation est le processus par lequel les espèces se forment. C'est une partie importante de la biologie de l'évolution.
Q : Que pensait Darwin de la formation des espèces ?
R : Darwin pensait que la plupart des espèces provenaient directement d'espèces préexistantes, ce qu'il appelait l'anagenèse ou "évolution phylétique".
Q : Comment la spéciation était-elle généralement perçue au 20e siècle ?
R : Au 20e siècle, les scientifiques pensaient que la plupart des espèces se formaient lorsque des espèces antérieures se divisaient, ce qu'on appelait la cladogenèse. On pensait également que cette scission était causée ou favorisée par des mécanismes d'isolement tels que la séparation physique.
Q : Comment les recherches récentes ont-elles changé notre compréhension de la spéciation ?
R : Des recherches récentes ont montré que l'hybridation entre espèces apparentées peut se produire et que des gènes peuvent être transférés par ces croisements. Cela signifie que l'isolement reproductif n'est pas toujours nécessaire pour définir une espèce et que l'allopatrie (séparation physique) n'est peut-être pas nécessaire pour que la spéciation ait lieu.
Q : Quel est le rapport entre la séparation physique et la formation de nouvelles espèces ?
R : La séparation physique entre des espèces autrefois coexistantes est considérée comme un facteur principal dans la formation de nouvelles espèces, illustré par de nombreux exemples discutés dans la littérature scientifique.
Articles liés
Auteur
AlegsaOnline.com Spéciation Leandro Alegsa
URL: https://fr.alegsaonline.com/art/92570
Sources
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833